The Undiscovered Country
By Robert Dorit
The statistics of death show leaps in modern life expectancy but fail to answer the question: Why do we die?
The statistics of death show leaps in modern life expectancy but fail to answer the question: Why do we die?

DOI: 10.1511/2007.67.398
I do not often picture my own death. When I do, however, the details vary. I can picture my death as being unexpected: a stroke, a heart attack, an auto accident. Some kind of testament to life's unpredictable character. But I prefer to picture the other sort of exit, one far in the future, where I have worn out my body without overstaying my welcome. Either way, I know that my story will end—I just don't know how.
We may be the only species that is aware of its own mortality. Yet despite death's central role in shaping human self-consciousness, mortality remains an elusive biological phenomenon. Fate and accident shape some ends, but these things aside, we cannot answer what seems like a straightforward question: Why do we die? The question, of course, is not really simple, nor does it yield a single answer. We will each die in our own way. But an answer collected from individual stories is not what we are after. We are, instead, seeking a more general explanation, rooted in material cause, which accounts for the patterns of human mortality. For now, we will leave out deaths that come from external causes—accidents or acts of violence—for they tell us little about the biological underpinnings of mortality.

© Private Collection/Giraudon/The Bridgeman Art Library
The pattern of death has changed through history. We can infer something about this pattern from fossils of early Homo sapiens. Judging by the condition of their skeletons and the extent of tooth wear, their life expectancy has been estimated at around 25 years. Tens of thousands of years later, as written records and gravestones become available, our ability to estimate life expectancy takes a major leap forward. The story told by these later records is dramatic: In the past 1,000 years, life expectancies and, presumably, their underlying causes have fundamentally changed. In the United States, for instance, the Social Security Administration has predicted life expectancies for the year 2050 will reach 77 years for men and 83 years for women.
So what are we to make of so drastic a change, a tripling of life expectancy at birth in 50 generations? It's a common misconception that a 30-year-old in the year 1000 would have been both old and rare. However, a 30-year life expectancy doesn't mean that at all. Many 50-year-olds walked the roads in the year 1000. Life expectancy at birth is an artificial construct, a statistical measure that averages survivorship across all age groups and that predicts how long an infant born today is likely to live given the death rates in effect today. That less-than-intuitive definition is worth pausing for, because its implications are profound. In effect, in the year 1000 your chances of making it to a ripe old age depended above all on surviving the first years of your life. If you turned 5, you were quite likely to turn 60. Life expectancy a millennium ago was determined primarily by rates of infant mortality. Over the next 800 years, and most particularly over the past 100 years, gains in life expectancy have come primarily from greater survival of infants and small children, often because of public sanitation and better treatment and prevention of infectious disease. A sad caveat: 27 of 136 countries in the world still have life expectancies at birth of under 50 years. For the half billion people in those countries, the gains in infant and child survival have yet to take deep root, or else have been offset by the dramatic increases in the mortality of young adults brought about by war and the HIV pandemic.
More recent increases in life expectancy in the developed world no longer stem from better infant and child survival. Increased life expectancy now comes about through two unrelated causes: a decrease in the number of children (which reduces the impact of child mortality on the calculation of life expectancy at birth) and an increase in the survival of older adults. Today, our life expectancy is greater not because we make it to adolescence, but because we are actually living longer, and the survivorship of older adults now has a far greater influence on the calculation of life expectancy.
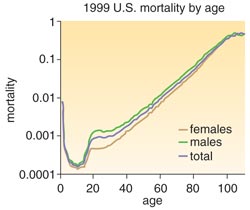
Given the many factors that enter into the calculation of life expectancy, however, this measure may not be the best way to understand death. Instead, we may want to look more closely at what demographers call age-specific measures, which quantify the probability of surviving a particular interval of life (usually a five-year span—ages zero to five, for example, or 40 to 45). We have known for a long time that surviving the first five years is a struggle, whereas the intervals between 5 and 30 are far less hazardous. We can also focus on the rate of mortality at a specific age, also known as the force of mortality.
Stephanie Freese
Much of our understanding of the force of mortality is rooted in the work of a brilliant 19th-century English mathematician, Benjamin Gompertz. Unable to enter the English university system because he was Jewish, Gompertz taught himself mathematics and was eventually elected into the Royal Society in 1819. In 1825, he published in the Philosophical Transactions of the Royal Society of London "On the Nature of the Function Expressive of the Law of Human Mortality, and on a New Mode of Determining the Value of Life Contingencies." This 70-page paper with its sexy title was Gompertz's effort to help underwriters calculate life-insurance rates. In preparing the paper, Gompertz had examined death records from several English towns and noticed a clear pattern for age-specific death rates. This pattern, now known as the Gompertz curve, shows a well-behaved, exponential increase in the force of mortality with age. He argued that death, seemingly shaped by so many chance events, nonetheless follows a simple and predictable path. And so simple a path could only arise from an equally simple, age-dependent, underlying force that operates throughout our lifetimes.
It took almost 150 years for Gompertz's curve to be seriously questioned. In part because of increased life expectancy in certain parts of the world, more individuals were living longer, providing sufficient data to examine the force of mortality in later years—data that Gompertz didn't have. The result of adding the long-lived to the analysis was startling: The force of mortality, which grows exponentially throughout most of one's lifetime, actually moderates after age 75. Equally unexpected was the discovery that the dramatic differences in age-specific mortality around the world virtually disappear after age 75. In effect, if you are born in Bolivia, your chance of seeing 75 is significantly lower than if you had been born in Canada. But if you make it to 75 in either place, your chance of seeing 90 is virtually the same. Past a certain age, the imprints of birth and circumstance seem to give way to universal forces and constraints.
Keep in mind that we have been speaking of age-specific rates of death, and not the cumulative probability of death, which, like it or not, always sums to 100 percent. In the end, we all die. But in the meantime, we seem to be succeeding in delaying the inevitable. For optimists and narcissists, the trend of increasing lifespan extends over the horizon. Conversely, demographers, biologists and politicians have asserted over the past century that human beings cannot possibly live beyond 70, or 80, or 85, or the nice round number of 100. Yet as they speak, the maximum age at death continues to climb in many parts of the world. For what it's worth, Jeanne Calment lived to be 122 years old, died in 1997, and remembered meeting the abrasive and ill-smelling Vincent Van Gogh when he came to live in her hometown of Arles, France. Although this trend of ever-lengthening lifespans cannot continue indefinitely (even for optimists and narcissists), the actual limit—and even the demonstrable existence of a limit—is a subject of continuing controversy among demographers and gerontologists.
This controversy arises in part because convincing statistics about centenarians remain hard to come by, increasing the temptation to extrapolate from isolated cases. Until recently, the number of people aged 95-110 was so small that reliable age-specific mortality rates for that age bracket were hard to estimate. People living past 100, moreover, might represent a small, but unique, subsample of the human population. Their age-specific mortality rates, buried for the first 80 or so years in the mass of data from the shorter-lived, may well have been different throughout their entire lives, only coming to dominate the statistics as the rest of the population drops off.
But for now let's leave aside the question of how long humans can live and concentrate instead on why most of us will not live to see the century mark. In 1990, the illustrious Russian biologist Zhores Medvedev sought to classify the more than 300 theories that had been put forth to account for this ineluctable reality. Many of these theories, compelling in their own right, likely capture some of the underpinnings of our mortality. Their sheer number, however, suggests as much about the complexity of the question as it does about the difficulty of thinking dispassionately about death. Scientists, after all, are people who are going to die too.
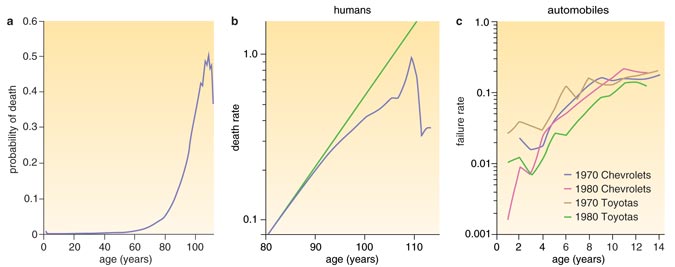
Data in (a) come from the National Center for Health Statistics, http://www.cdc.gov/nchs/deaths.htm. Figures adapted by Stephanie Freese from T. C. Goldsmith, http://www.azinet.com/aging/Aging_Book.html, and Vaupel et al.
Theories of death are also deeply embedded in their culture of origin. Let's consider, for instance, the class of mechanical explanations of death. These hypotheses invite us to accept the metaphor of the human body as a machine with many parts that must work in concert. In this metaphor, we die either because a key component gives way (the catastrophic failure model) or because several interacting parts begin to wear out and no longer work properly (the systems failure model). These metaphors of failing machinery compel, but data supporting them are harder to obtain. Catastrophic failures, like heart attack and stroke, are major killers in the developed world, particularly in the United States. Many of these deaths, however, are preventable and linked to a sedentary lifestyle, and not to an inherent age-dependent failure of a particular organ system.
The systems-based explanation argues that complex machinery fails because many things go wrong. According to this model, as with your Toyota, so too with your body: The force-of-mortality curves for automobiles (called failure-rate curves) complied by demographers James W. Vaupel and Cynthia R. Owens at Duke University bear an uncanny resemblance to their human counterparts. Both human and automobile curves show an exponential increase in the force of mortality that tapers off in later years—the same violation of the Gompertz Law we spoke of earlier. Surprisingly, mortality in the early years is eerily similar in people and automobiles: Defects in manufacture (machines) or development (organisms) reveal themselves early on. But if you (or your car) make it past the first year, you're likely to motor along for a good period thereafter. The patterns are tantalizing in their similarity and may reveal some fundamental features of failure in complex systems. Such systems, whether we speak of Toyotas or biologists, are characterized by redundancy brought about by engineering (in the first case) or evolution (in the second). But, as University of Chicago gerontologists Leonid Gavrilov and Natalia Gavrilova have argued in their influential bookThe Biology of Life Span: A Quantitative Approach, the very redundancy that permits complex systems to endure a constant rain of light damage also allows such damage to accumulate, resulting in aging and eventual failure. Surprisingly, the higher the level of redundancy, the faster the force of mortality accelerates with age. The irony is that the redundant mechanisms that ensure survival during our early years are the same ones that speed our twilight exit.
For two centuries, biology has profited from Rene Descartes's conception of the human body as a machine, subject to, and more importantly, explainable by, material laws. Compelling as the Cartesian metaphor of the body-as-machine might be, it may, however, conceal some important differences between my automobile and me. Repair, for instance, is a process central to survival. I depend on my excellent mechanic, an agent external to my automobile, to keep my car on the road. In contrast, bodies, for the most part, repair themselves: More than 100 genes in our genome are devoted to detecting and repairing DNA damage.
This notion of the body as a self-policing, self-correcting system has spawned a second class of death explanations, those related to maintenance. Although the human body can detect and remedy a startling array of defects, this capacity to self-renew declines with age. Even the raw materials for repair—those stem cells recruited to replace exhausted counterparts—become depleted. The damage to our bodies, meanwhile, continues unabated throughout our lives. Much like the inventive mechanics in Cuba who keep Edsels and Cadillacs from the '50s running without spare parts, the body struggles to patch up broken functions as replacement cells decline. But it is, ultimately, a losing battle. In a final ironic twist, the repair process, like the existence of redundancies, becomes a source of trouble as we get older. The inflammation that accompanies injury and repair is itself a source of further damage. Aging and death may come about when we can no longer keep up with the maintenance required to stay alive.
A third class of evolutionary explanations for aging and death suggests that natural selection focuses largely on the young and the fertile and is indifferent to those past their reproductive years. In the years prior to reproduction, for instance, natural selection appears to suppress many genes that underlie disease. After reproduction, however, when such genes have comparatively minor effects on an organism's reproductive success (though not on its health), they are free to exert any malign influence. This scenario may explain why so many diseases, notably cancer and cardiovascular disease, increase precipitously in the fifth decade of our lives. Mutations that benefit the young—even if they cause problems later in life—will always gain traction.
Further support for an evolutionary approach to understanding death comes from the demonstration that fruit flies prevented from reproducing until late in life evolve significantly longer lifespans. By delaying reproduction, the experiment keeps natural selection in play for a longer amount of time, favoring genetic changes whose deleterious dark side is also delayed. In just a few generations, average lifespan increases. Nevertheless, harmful mutations eventually, inevitably accumulate into insurmountable, irreparable damage. These models lead to one conclusion: Aging and death occur when natural selection takes its eye off the ball in the later years.
A newer set of theories, prompted in part by our predilection for molecular explanation, have recently come on the scene. Such arguments—we'll call them fatalistic theories—suggest that a series of clocks tick away at the cellular level, marking our remaining time. They argue that evolution has programmed organisms to know when it is time to die. Critical to this argument is the observation that in a lab dish, most mammalian cells divide about 50 times and then die. We now know that chromosomes, those densely packed spools of DNA inside each cell, get shorter every time they're copied. Past a certain point, this chromosomal shortening kills the cell.
But we still can't connect the mechanisms of cell death with the mechanisms of our death (or, for that matter, the death of any other multicellular organism). Once again, the body is not just a collection of cells that behave independently. Our ability to repair and regenerate, to conscript circulating stem cells—whose chromosomes don't shorten when copied—into regenerating tissues and organs significantly complicates these hypotheses of aging and death.
More recently, some weight has been given to the idea that longevity is simply the result of certain trade-offs. Experiments showing that very low caloric intake (starvation) significantly increases lifespan suggest that there is a tradeoff between lifespan and active metabolism (which produces damaging chemical waste). Many studies in other species show a negative correlation between reproduction and longevity, suggesting that energy spent in reproduction would otherwise be spent in living a longer life. Some find in these trade-offs a guide for living. To me, their emphasis on the longevity benefits of an ascetic lifestyle sound slightly puritanical.
Ultimately, what does it suggest to have so many competing theories of death? I would argue it is the hallmark of a vibrant field, one in its early phases. The many explanations jostle and compete, driven by our shared belief that death may be inevitable, but it is not incomprehensible. The identification of genes involved in determining lifespan, the demonstration that lifestyle and environment can delay death, the technologies that extend life—all fuel our fascination with the topic. But the aging population in the developed world (including its scientists) also propels the field. In the end, however, if our real motivation is to loosen death's grip on humanity, we need to go beyond a material understanding of the biology of aging, lifespan and death. Most of humanity still does not die of old age, "sans teeth, sans eyes, sans taste, sans everything," as Jaques laments in As You Like It. Instead, most of humanity dies young, of preventable causes brought about by poverty and unequal access to health care. Any account of the causes of death that focuses solely on biology without considering the social and economic settings in which our biology plays out is at best incomplete, and at worse, misguided. Over my lifetime, cheating death may not be about tricking the limits of our biology, but instead about changing how we live our lives.
Click "American Scientist" to access home page
American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.