The Rise and Fall of Rudist Reefs
By Claudia Christina Johnson
Reefs of the dinosaur era were dominated not by corals but by odd mollusks, which died off at the end of the Cretaceous from causes yet to be discovered
Reefs of the dinosaur era were dominated not by corals but by odd mollusks, which died off at the end of the Cretaceous from causes yet to be discovered

DOI: 10.1511/2002.10.148
From the mountains of Europe to the deserts of Mexico, one finds geologic formations that contain ancient marine fossils so peculiar in shape that local folk once imagined them to be horns shed by sheep or goats. Some farmers built impressive walls and fences with these odd stones. Many decades ago, they realized that their building materials were in fact the shells of extinct marine organisms.
But few of those people could have grasped just how apropos it was to be arranging those particular fossils into imposing barriers. After all, without some training in geology, who would know the ancient animal's true notoriety in the annals of the former world: that of being the only mollusk ever to construct massive reefs.

Photograph courtesy of Carl Koch.
That understanding, of course, took a long time to develop. When the French naturalist Jean-Baptiste Lamarck examined these fossils in 1819, he coined a term for them—rudistae, or "rudists," from the Latin rudis (meaning "rude" or "raw"), or possibly from rudus (denoting rubble or broken stone). Lamarck may have suspected that his rudists once occupied shallow seas, but he would not have guessed that they might have made a tasty dinner. After all, they didn't look at all like their distant cousins, the scallops, mussels and oysters that he probably enjoyed eating.
For years, scientists tried to shoe-horn rudists into various phyla before placing them ultimately in Mollusca. It was longer still before all rudists were moved from Cephalopoda to Bivalvia, a 42,000-species class that evolved more than 500 million years ago.
Today, at least 10,000 bivalve species live in freshwater lakes and streams, in estuaries and bays, and in shallow coastal and deep-ocean environments. Because bivalves have been successful at invading and flourishing in habitats spanning a wide range of latitudes and temperatures, it's not surprising to find that they also live in tropical reef communities, which are made up of an ecologically complex mixture of corals, sponges, algae, snails, fish and many other groups.
Defined simply, reefs are masses of rock lying at or near the surface of the water. As such, they are barriers to navigation; the most spectacular examples lie off the coasts of Australia and Belize. A more complex definition considers the ecology and the physical and chemical attributes of the surrounding water and sediment—the reef as an integrated community of bottom-dwelling organisms that secrete a rigid skeletal framework, forming a wave-resistant structure.
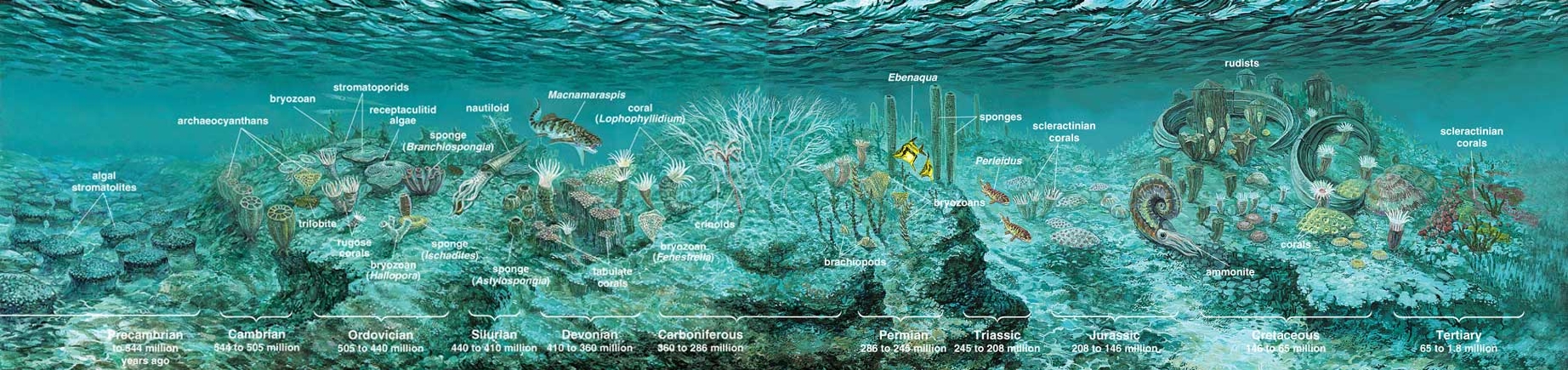
Although people now refer to tropical ecosystems of this sort simply as coral reefs (in deference to the chief skeletal builder today), reefs were not always constructed of corals. Paleontologists have identified reef ecosystems in strata dating back at least to the start of the Paleozoic, perhaps earlier. These ancient reefs were dominated by animals with unfamiliar names such as archaeocyathids and stromatoporoids, and by the more familiar sponges and extinct corals such as rugosans and tabulates. The modern types of corals are new to the scene, having evolved recently, geologically speaking, just before the arrival of the rudists.
The oldest rudists are found in late Jurassic rocks in France, ones formed about 160 million years ago. These creatures appear in Caribbean formations that are several million years younger. Rudists probably originated in Europe and made their way across an ocean that geologists call Tethys, a former seaway that extended from what are now the Himalayas to Mexico. Rudists thrived through most of the warm Cretaceous period, a time when the Pacific Ocean was wider than today, and North and South America had not yet joined.
Rudists spread throughout the tropical seas, whereas only rare individuals and isolated clusters survived at higher latitudes. Rudists were more common in the Northern Hemisphere, a fact that can be attributed both to their northerly origination and to the dominant east-west circulation pattern across the Tethys sea.
Direct evidence for the early dispersal of rudists is scarce, but paleontologists infer that the larvae of rudists, like those of many modern bivalves, were carried along with plankton in surface currents, giving rudists their cosmopolitan distribution. Expanding on the ideas of previous investigators, I made use of this phenomenon and was able to chart Cretaceous circulation patterns for the Gulf of Mexico and the Caribbean by plotting rudist distributions on maps of the former geography (ones that take account of plate-tectonic movements) and noting the presence or absence of various species. High statistical correlations from place to place indicated the location of ancient dispersal routes.
Although rudist larvae could move with the currents, it seems clear that once their passage across the slowly expanding Atlantic was complete, time and distance isolated them from their European forebears. This separation drove the evolution of different species and reef structures on the two sides of the Atlantic.
Rudists were not, however, the sole reef builders at the time. In the calm, shallow waters of the low-latitude Cretaceous, the rudists initially shared space with scleractinian corals (the type that dominates tropical reefs today). But by the middle Cretaceous, about 100 million years ago, rudists dominated the carbonate platforms (shallow areas largely covered by the calcium carbonate shells of marine creatures), taking over much of the space formerly occupied by corals. This ecologic shift happened several million years earlier in the Caribbean than in the Mediterranean. Geologists do not yet have sufficient material from the Pacific to evaluate whether the same changeover took place there. Still, it is clear that over the Cretaceous, rudists eventually outnumbered corals in most parts of the world.
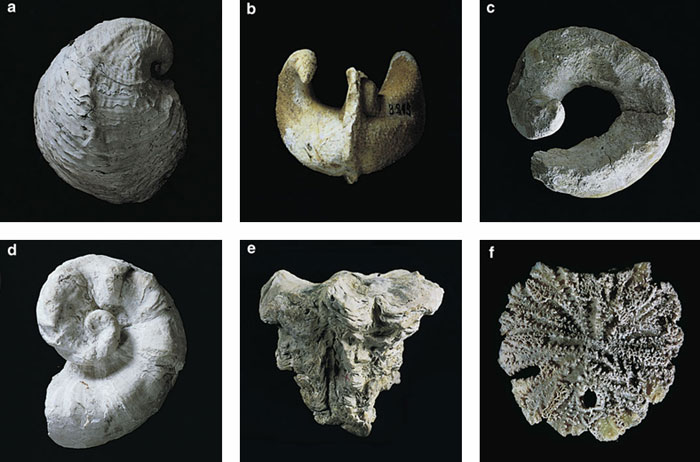
Rudists were in many ways perfectly adapted for the tumultuous life on a reef. Like other bivalves, they were protected by two hard calcareous shells, or valves, attached to each other along a hinge line, allowing them to open and close.
D. W. Miller
The rudists constructed their durable homes by extracting calcium and bicarbonate from seawater and precipitating multilayered shells made of calcium carbonate. Their shells were divided internally to provide a space for the body as well as various accessory cavities and canals. One valve usually attached to a hard surface on the sea bottom and had teeth and sockets along the hinge, whereas the opposing valve had the opposite dental configuration. This arrangement held the two valves firmly in place after they closed.
Opening and closing its shell is an essential function for a modern bivalve. These actions enable the animal to deploy its siphons (special tubes for feeding), to extend its foot (an appendage for burrowing) and to seal itself off from predators. Like their modern counterparts, most rudists used ligaments to open their valves and muscles attached to the inside of the shell to close them. But from this simple scheme emerged many variations.
Some rudists lacked connective ligaments and may have had to pop open their valves by elevating the pressure inside the shell. Some rudist fossils show what are thought to be siphonal bands on the side of the shell. These bands represent the positions where the siphons would have rested; the siphons themselves are not preserved. So questions linger about how these creatures derived nourishment. For example, whether they depended on suspension feeding or filter feeding (and the size of the particles they ate) remains open to debate. Paleontologists also ponder whether the rudists lived in, above or partly buried in the sediment. (Most modern bivalves with siphons live in the sediment; those without live on top of the sediment.) But the answer to this question about rudist lifestyles may have depended on the shape and size of the shell, which differed widely between species, rather than on the presence or absence of siphons.

D. W. Miller
The variation in rudist size and shape was indeed extreme. Measurements of fossil valves range from a few centimeters to a meter—among the largest bivalves ever recorded. Whereas the two concave shells of a typical cockle or clam mirror each other in size and shape, the rudists presented a dramatic evolutionary departure from this symmetrical design.
Some rudist valves were coiled, some curved. Others were conical or cylindrical, and some were thin and flat. In some cases, a large conical lower valve was hinged to a small flat upper valve; in other types a small conical lower valve was attached to a large coiled upper valve. It is clear from their geometry that some of these forms would have needed to prop themselves up using the surrounding sand or mud. The amount of sediment required to stabilize the shells would have varied according to shape, and it is likely that not all types needed such bracing. Shells often grew close to one another, forming networks of mutual support.
Such congregations could become large—quite large. Indeed, some reefs of rudist shells ran for hundreds of kilometers along the edge of carbonate platforms. These reef structures withstood the normal pounding of the waves but were destroyed by hurricanes and other large storms. Rudists repeatedly regrouped after such tempests, in some cases building on rubble that had fallen down the steep reef slope, so as to form structures that were hundreds of meters tall. Such thick, porous accumulations of broken rudist debris later became valuable oil reservoirs in the Gulf of Mexico and the Middle East.
Large piles of rudists are, however, rare in the Mediterranean region. There rudist formations contain cylindrical valves packed tightly together, with many of them touching and some surrounded by sediment. Coral patches, large and small, cropped up among these rudists. Algae, sponges and stromatoporoids also lived with them and increased biologic diversity on these Cretaceous reefs. The degree of biodiversity, however, was far less than that which we are accustomed to seeing on tropical reefs today. The lack of diversity was due, at least in part, to the especially warm environments that prevailed in the Cretaceous.
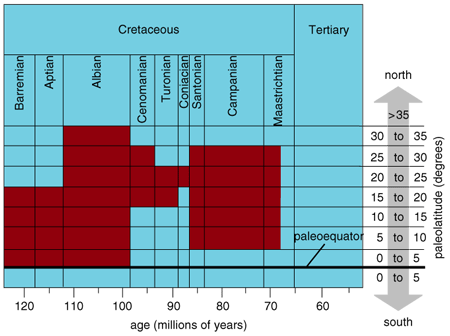
During the Cretaceous, the Earth was about 6 to 14 degrees (Celsius) warmer than it is today. Such balmy conditions made for a very different world. There was, for example, no permanent ice at high latitudes. And the temperature of tropical surface waters of the open ocean was warmer than today, perhaps reaching 30 to 32 degrees. In the shallower waters where rudists lived, temperatures may have been higher still, although it is difficult to gauge the temperature of coastal waters using the standard technique (the geochemical analysis of the shells of ubiquitous marine creatures called foraminifers). In regions with restricted circulation—for example, on broad carbonate shelves or on reefs near land—salinity may have been high, too. Associated evaporites (rocks made up of minerals that form from evaporating seawater) testify that rudist habitat during the Cretaceous period was both warmer and more saline than are most tropical waters today.
Obviously, some rudists must have thrived in these hot, salty conditions. But the even higher temperatures then prevailing near the ancient equator may have had the opposite effect on the carbonate platforms in the Pacific, leading to the demise of those carbonate platforms and, by implication, to rudists in that part of the world.
Working with a computer model of the ocean and atmosphere, several investigators (including Eric Barron from Pennsylvania State University) have tried to mount simulations that show an average global temperature in line with known Cretaceous conditions. To do so, they needed to boost atmospheric carbon dioxide levels in their model, yet they found that high atmospheric carbon dioxide—four times the present-day value—translated to overheated tropics and overly cool polar regions. The tropics would display normal levels in the model only when the simulated oceans were forced to transport heat poleward, which in turn warmed the temperate zones and polar regions.
This modeling of Cretaceous climate suggests that the ocean must have carried enormous amounts of heat away from the equator, as it does today. Working with Barron and other investigators, Erle Kauffman (of Indiana University) and I surmised that at specific times in the mid-Cretaceous so much heat was transported away from the tropics that the preferred environment for rudists collapsed—as did rudist diversity. This hypothesis neatly explains why tropical conditions in the normally warm Cretaceous oceans were occasionally restricted to a narrow range of latitudes.
This scenario, which depends critically on the ocean's capacity to transport heat, fits in well with what we know about the Atlantic today, where sea-surface temperatures peak north of the equator and the highest salinity surface water is found at 30 degrees North. These conditions reflect the fact that surface currents carry a considerable amount of heat northward from the equator, and when these warm surface waters evaporate, the top layers of the ocean become especially salty. The close match between the model results and empirical data provides a compelling case for why rudist diversity waxed and waned from the middle to the late Cretaceous. This work does not, however, offer an easy answer to the question of why the rudists died off at the end of the Cretaceous.
Reconstructing ecological conditions surrounding the extinction of the rudists is a work in progress, but geologists know some facts for certain right now. Kauffman and I calculated that rudist reefs disappeared from the Caribbean either 3 or 1.5 million years before the end of the Cretaceous, when a host of other Mesozoic creatures suddenly went extinct, including, most famously, the dinosaurs. Possible causes for the sudden demise of Caribbean reefs include a general cooling in tropical temperatures and the dwindling of reef space that accompanied a drop in sea level.
Jean Philip and his colleagues at the Université de Provence reported that the rudists went extinct in western Europe and in regions surrounding the Adriatic Sea about 0.5 million years before the very end of the Cretaceous. Elsewhere, such as the central-eastern Mediterranean and Middle East areas, the extinction of the rudists seems to have taken place somewhat earlier. Some investigators have suggested that a drop in sea level, which removed the habitats for rudists, caused their extinction in this area.
David Schneider
Yet it remains rather mysterious why the rudists went extinct before the disaster that ended the Cretaceous period—one sparked by a meteorite that struck the Yucatan Peninsula some 65 million years ago. That great collision is thought to have launched vast quantities of dust into the stratosphere, where it lingered, blocking out sunlight for months or even years. The subsequent shut-down of photosynthesis, as well as a host of other impact-generated ecological disturbances led many groups (including the dinosaurs) to extinction. It took perhaps hundreds of thousands of years for the environment to recover to the point that large animals could once again roam the continents. It took a much longer time—7 to 10 million years—for the scleractinian corals and associated biota to reorganize into the tropical reef ecosystems seen across the globe today.
Clearly, the end of the Cretaceous was a time of immense stress for many creatures. About two-thirds of the marine species were lost during what has become recognized as a "mass extinction." Some environmentalists warn that the natural world today faces similar dangers. Tropical reefs in particular are vulnerable to the effects of global warming, because excessive heat can cause so-called coral bleaching and death. What is more, Joanie Kleypass of the National Center for Atmospheric Research and her colleagues have recently shown that as atmospheric carbon dioxide levels rise, oceanic chemistry will shift enough to threaten corals as well.
So it seems entirely possible that tropical corals, and the distinctive biological communities they support, might suffer in the future. Should corals die off entirely, other marine organisms will surely take their place, although just what sort of plant or animal will dominate future reefs is uncertain. If the heat-loving rudists were still alive today, they might well be the most fit to take over. Then again, if rudists were alive today, they might be endangered from being on too many menus.
Click "American Scientist" to access home page

American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.