Relative Pitch and the Song of Black-Capped Chickadees
By Ron Weisman, Laurene Ratcliffe
Chickadees, like people, have a strong sense of relative pitch. These birds use skillful, precise pitch changes to advertise their quality and attract mates
Chickadees, like people, have a strong sense of relative pitch. These birds use skillful, precise pitch changes to advertise their quality and attract mates

DOI: 10.1511/2004.50.532
More than 2,000 years ago, the acerbic philosopher Marcus Tullius Cicero observed that Roman songbirds compose more excellent melodies than any musician. He certainly doesn't stand alone in history on that count; it is a nearly universal human experience to find joy and wonder in birdsong—and to compare the songs to human music.
People have been transcribing melodies of birds into the notation of music since at least the 18th century; Vivaldi's Goldfinch concerto and Handel's Cuckoo and the Nightingale organ concerto include musical notation for birdsongs. In the early 1900s, the New England naturalist and composer F. Schuyler Mathews presented the songs of many North American birds in musical notation in his Field Book of Wild Birds and Their Music in order to help readers identify species common in the eastern United States.
Later biologists did not share Mathews's enthusiasm for musical descriptions of birdsongs. Donald Borror, a master bioacoustician and field biologist, found many of the song descriptions inadequate by modern standards, as he wrote in his foreword to the 1967 reprinting of Mathews's book. Borror acknowledged that Mathews lacked modern electronic equipment and that his primary interest was in the musical content of birdsongs. Today, however, Mathews's approach seems dated and quaint. Musical notation is simply unable to provide the detailed, accurate and reproducible descriptions required in modern bioacoustical analyses of vocal communication among songbirds. Mathews's approach helps musically trained people recognize birdsongs, but it fails to objectively describe birdsongs and calls. For bioacousticians, accurate observations are a crucial first step in analyzing the role of songs in the life of a species.

Photograph courtesy of Dan Mennill.
Modern ornithologists believe that songbirds first appeared some 40 million to 50 million years before human beings. That people derive pleasure from birdsongs and recognize their musical features suggests that despite the vast evolutionary gulf between birds and mammals, songbirds and humans share some common auditory perceptual abilities. In this article, we focus on relative pitch, the ability to recognize relationships between acoustic frequencies. We review studies suggesting that humans and songbirds share the ability to perceive relative pitch changes and exploring the evolutionary functions of birds' perception and use of relative pitch.
Relative pitch is the ability to produce or recognize a note based on a reference note. The ability to hum a simple tune when given a set note with which to start, as most people can, shows relative pitch in action. A sense of relative pitch is common among non-musicians even if they never use the term, but musical training develops this knack. Trained singers have an advanced appreciation of pitch change: They can produce precise relative frequency changes from one note to the next in a melody.
To compare the appreciation for relative pitch among humans and songbirds, it is necessary to understand how musicians and bioacousticians measure pitch change. Musicians describe the change between any two notes on the musical scale as an interval. The interval between any two adjacent notes is defined as a semitone. The ratio between the frequencies (measured in cycles per second, or hertz [Hz]) of the higher and lower of two notes separated by a semitone is always 1.059, regardless of where the two notes are located on the scale. Musicians speak of semitones and intervals; bioacousticians use the terms frequencies and frequency ratios.
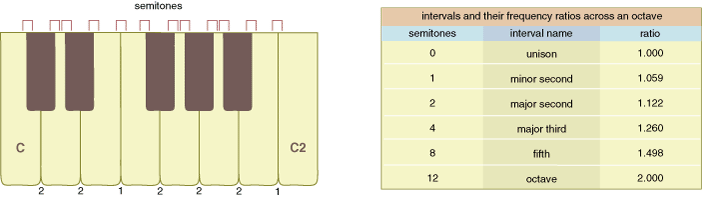
Illustration by Emma Skurnick.
Pianos and keyboards easily illustrate the discrete intervals between notes. For example, hitting the E note in the fourth octave (E4, 330 Hz), then counting down four keys (where both black and white keys are counted) and hitting the C note in the same octave (C4, 262 Hz) produces a relative pitch change of four semitones, as measured by musicians, and a frequency ratio of 1.26, as measured by bioacousticians. One can easily transpose a four-semitone pitch change on a piano. For example, moving up the keyboard and hitting the E note in the fifth octave (E5, 659 Hz) and the C note in the same octave (C5, 523 Hz) produces the same relative pitch change as in the fourth octave: four semitones or a frequency ratio of 1.26. Although they use different notations, musicians and bioacousticians agree on the constancy of pitch change across the keys. If one measured the frequencies produced by hitting E4 and then C4 on each of 20 pianos, the average ratio should be 1.26 with a very small standard deviation among the pianos. The small standard deviation helps illustrate that pitch changes in music are discrete.
Keyed instruments, such as pianos, are constrained to produce discrete pitches, whereas keyless instruments, such as violins and human voices, can produce pitches that vary continuously. In practice, however, trained violinists and singers produce discrete pitch changes analogous to a piano's keys. Put another way, although the human voice can vary in pitch continuously from one frequency to another, that is not the way humans use their voices or their musical instruments. People produce pitch changes with low variability about their frequency ratios.
Humans recognize pitch changes in discrete intervals and are good at identifying notes relative to an external reference pitch—in other words, relative pitch. Most people easily recognize a melody transposed to a different key, or starting frequency, because the song maintains the same intervals, or frequency ratios, among the notes. Trained musicians, of course, are better than others at identifying intervals and transposed melodies, but practically everyone has some sense of relative pitch in simple melodies. For example, people instantly recognize the song "Happy Birthday" regardless of the key in which it's sung.
Human musicians have not artificially adopted common scales to coordinate their musical performances; people, whether musicians or not, produce and perceive discrete, constant frequency ratios across different renditions of the same melody. In comparing relative pitch perceptions across species, we use low variability about mean frequency ratios in the same song across singers to provide evidence that songbirds, like people, have a strong sense of relative pitch.
Songbirds belong to the class Aves, which includes all birds, the order Passeriformes, consisting of perching birds, and the suborder Oscines, the true songbirds. The true songbirds are distinguished from other perching birds by the anatomy of their vocal organ, the syrinx, and by the fact that they need to learn their songs while juveniles from adult members of the same species. Oscines are an evolutionary success, comprising about half of the more than 9,000 species of birds and filling niches in almost every environment on the planet. Because songbirds are so diverse in appearance and behavior, no single species can represent the whole group. Our studies ask not whether all species of songbirds use relative pitch to produce and perceive songs but whether at least some Oscines do so in order to identify the songs of their own species. We have concentrated our studies in particular on forest-dwelling songbirds such as the black-capped chickadee, Poecile atricapillus (formerly Parus atricapillus), because the songs of forest birds contain pure tones—that is, tones of narrow frequencies—which are relatively easy to measure and reproduce in perception experiments, both in the laboratory and in the field.
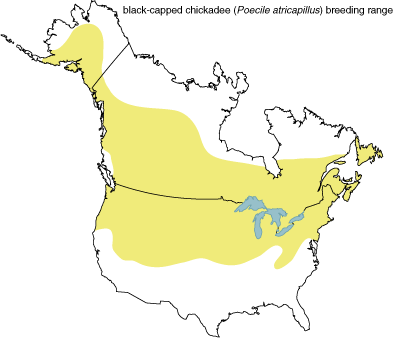
Illustration by Emma Skurnick.
Black-capped chickadees are familiar and abundant year-round in the northern two-thirds of the United States and in much of Canada. In his 100-year-old field book of songbird music, Mathews included black-capped chickadees. True to his description, these chickadees are acrobatic, noisy and highly social birds. Their name derives from their call, which sounds like "chick-a-dee." This call is given by male and female, young and old, to keep in touch with members of the flock. The chick-a-dee call is complex, and some of the notes include "buzzy" harmonics. During the breeding season and less frequently throughout the year, males sing the "fee-bee" song, used to hold territory and to attract and arouse females. We focus on the fee-bee song in this article.
Mathews observed that the male chickadee's fee-bee song consists of two or three clearly whistled notes. The first note, "fee," is sung at a higher pitch than the second one, "bee." Sometimes, he noted, the bee note appears to be sung as two separate notes at the same pitch. Mathews suggested that chickadees separate the fee and bee notes in their songs by a clearly perceptible pitch interval. Recall that musical intervals are simply precise discrete frequency ratios. About 70 years later, Steward Hulse of The Johns Hopkins University restated Mathews's hypothesis in modern bioacoustic terms when he suggested that chickadees might hold the ratio between the frequencies of their fee and bee notes constant across birds.
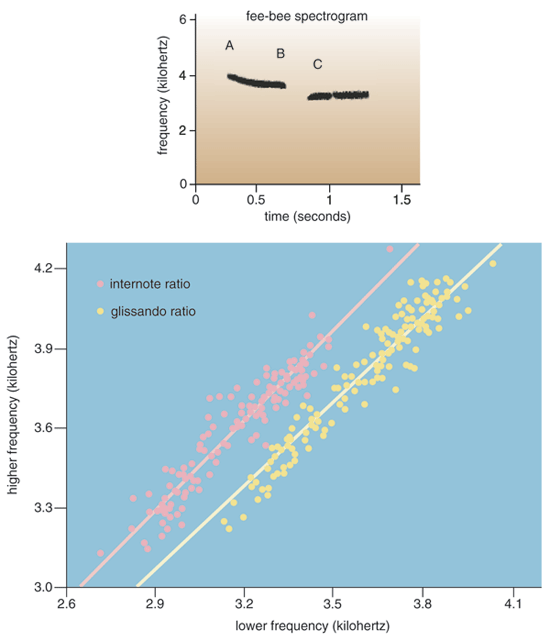
To test Mathews's and Hulse's hypothesis, we, along with Ingrid Johnsrude, now at Queen's University, and Andy Hurley, now at the University of Lethbridge, conducted a large bioacoustic analysis of several songs from each of 154 male black-capped chickadees singing on their territories in eastern Ontario. We found that, indeed, the internote ratio between the frequencies of an individual chickadee's fee and bee notes varied by less than 2 percent about a mean value of 1.134 (just slightly more than two semitones). Also, the ratio between the fee and bee notes varied less than 2 percent across our sample of chickadees. Our spectrograms showed that the fee note is not constant in pitch but instead sweeps downward in frequency in what musicians call a glissando. The frequency ratio of the decline in the pitch from the start of the fee note until its end was 1.056 (just less than a semitone) with approximately 1 percent variability among the renditions either by an individual bird or among birds.
The spectrograms show that male black-capped chickadees' songs contain not one but two relative pitch cues: the decline in frequency from the start to the end of the fee note—the glissando ratio—and the decline from the end of the fee note to the beginning of the bee note—the internote ratio. Quite unexpectedly, we found that individual males that were brought into the laboratory, isolated and recorded briefly occasionally sang their fee-bees on different starting pitches. Additional field observations by Brad Hill, then a graduate student, and Ross Lein, at the University of Calgary, confirmed these findings. With our collaborators, Andy Horn and Marty Leonard, now both at Dalhousie University, and Scott MacDougall-Shackleton, now at the University of Western Ontario, we measured many more pitch-shifted songs from individually tagged males during their extended and persistent dawn chorus. The chickadees surprised us. In the first dawn recording session, each bird sang his fee-bee song at a wide range of frequencies, changing frequency on average every 40 songs. Over subsequent sessions, individual males filled in the gaps between frequencies within the range we heard.
These bioacoustical measurements taught us some astonishing facts about song production in black-capped chickadees. Individual chickadees do not use discrete keys to place the starting frequencies of their song but instead vary their songs continuously across the range of frequencies used by members of the species. In other words, when chickadees change frequencies from the start to the end of the fee note and from the end of the fee note to the start of the bee note, their pitch changes are very precise, but their starting pitch varies. In musical terms, black-capped chickadees transpose the sequence of relative pitch changes typical of the song of their species across a wide range of pitches. This combination of musical abilities is rare in birds. Recordings of other songbirds—in particular, white-throated sparrows (Zonotrichia albicollis) and veeries (Catharus fuscescens)—show that individual birds do not change the starting frequencies of their songs from rendition to rendition. Constant note frequencies give other members of the species a way to identify individual birds. At the same time, a constant frequency ratio in the song of the species helps identify the species.
Humans may listen to birdsongs for solace and entertainment, but among songbirds, including black-capped chickadees, males listen to one another to identify potential rivals for territory, and females listen to find potential mates who possess territories. During the breeding season, male chickadees defend territories of, on average, about 5 hectares from rival males and attract females with whom they reproduce and rear young. If a rival male sings in another's territory, the male already established there drives out the intruder by singing vigorously, approaching the trespasser and, if the rival persists, physically attacking him.
For more than four decades, to discover how songbirds communicate, biologists have conducted experiments playing back songs. Recordings can simulate the intrusion of a rival male on or near another's established territory. Skillfully recorded and reproduced songs of the species elicit forceful territorial defense, whereas playing back songs with altered acoustic features may provoke a weaker territorial response, depending on the importance of the altered features to the potency of the song.

Illustration by Emma Skurnick. Adapted from a photograph by Dan Mennill.
We wondered whether accuracy in relative pitch is important to the potency of chickadee songs. In several studies, we played back normal fee-bee songs and a variety of fee-bee songs with altered frequency ratios to male chickadees on their territories. One of us (Ratcliffe), along with Scott MacDougall-Shackleton and Dan Weary, now at the University of British Columbia, looked at the roles of both the glissando and the internote frequency ratio in eliciting territorial defense. Territorial male chickadees in their native environment heard a normal song and altered songs lacking the glissando element. One altered song included a fee note flattened to the start frequency of a normal fee note, which increased the internote ratio relative to the bee note. A second altered song presented a fee note flattened to the end frequency of a normal fee note, which therefore left the fee-bee internote ratio unchanged compared with a normal song. Male chickadees flew closer to the speaker on more occasions and called more often to a normal song than to the playback of either altered song. The two altered songs provoked about equal territorial defense. The first altered song had abnormal glissando and abnormal internote frequency ratios, yet the male chickadees responded no less to it than to the second altered song, which had only an abnormal glissando frequency ratio.
Our results puzzled us. We could not understand why male chickadees produce accurate discrete pitch changes in their songs only to ignore the vastly inaccurate internote pitch changes they heard in playback experiments. We were skeptical of this conclusion in part because other songbirds—in particular male white-throated sparrows and veeries—are known to produce precise, predictable pitch changes in their songs, and playback experiments that altered these ratios reduced territorial defense in both species. We continued to seek reduced territorial responses in black-capped chickadees to songs with altered internote frequency ratios. In one series of experiments, we digitally copied and spliced together fee and bee notes from several chickadees to form songs with normal internote ratios and songs with ratios as disparate from normal as 1.00 (equivalent notes) and 1.29 (between four and five semitones apart). Judging by several behavioral measures, male chickadees persisted in defending their territories equally strongly against all of these songs, whether the internote ratio was normal or altered. Taken together, these field experiments have led us, finally, to conclude that the glissando frequency ratio is far more important to territorial defense by male black-capped chickadees than is the internote ratio.

Illustration by Emma Skurnick. Adapted from a photograph by Dan Mennill.
But what about female responses? Among most songbirds, chickadees included, females respond weakly or not at all to simulated territorial intrusions. But female choice is a powerful evolutionary force, and we decided to address whether female chickadees care about the pitch changes sung by potential mates. One of us (Ratcliffe) and Ken Otter, now at the University of Northern British Columbia, brought female black-capped chickadees into breeding condition in the laboratory by artificially extending their day length and administering the hormone estradiol. When female songbirds are in breeding condition, the songs of males from their species elicit a distinctive copulation-solicitation display. We played the receptive females two normal songs, a song with an increased internote ratio and a song with an altered glissando. Female chickadees displayed longer to both normal songs than to the song with an altered internote ratio, and they displayed more often to the normal songs than to the one with an altered glissando. In other words, the females chickadees appeared to discriminate sexually in favor of normal songs over songs with either glissandos or internote ratios altered. The surprising implication of our playback experiments is that although males need to produce a precise internote pitch change to attract and stimulate females, the males themselves seem not to care at all about whether their territorial rivals produce wildly inaccurate internote ratios.
Like human musicians, songbirds learn their songs; oscines do not produce precise frequency ratios in their songs by some feat of gene transcription. Most humans, of course, have some innate musical ability and can sing and recognize at least some melodies, and trained musicians clearly learn to perfect their relative-pitch skills. Such observations led us to consider whether black-capped chickadees learn to produce the precise, discrete pitch changes in their fee-bee songs. We have reared male chickadees isolated from adults on several occasions and have examined spectrographic records from these birds as well as from isolated birds reared by Steve Nowicki at Duke University, along with his students Melissa Hughes, now at the College of Charleston, and Bernard Lohr, now at the University of Maryland. Unlike chickadees reared normally, isolated males sing quavering, low-volume notes and seldom produce consistent pitch changes between notes.
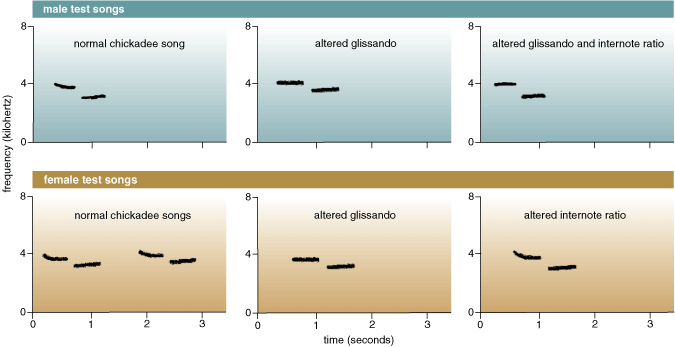
Illustration by Emma Skurnick and Stephanie Freese.
Some songbirds can learn the song of their species from tape recordings, but many species, including chickadees, learn best when they interact with live adult males. Of more than a dozen male black-capped chickadees reared with taped tutors by one of us (Ratcliffe) with Scott MacDougall-Shackleton, and by Don Kroodsma and his colleagues at the University of Massachusetts, only three birds learned good approximations of taped fee-bee songs, and only one bird shifted the pitch in his songs. The remaining birds sang several whistled notes and sometimes changed pitch dramatically mid-note.
Illustration by Emma Skurnick. Adapted from a photograph by Chris Sturdy and Isabelle Charrier.
From these studies of isolated and tape-tutored males, we concluded that young male chickadees acquire their precise control over the production of relative pitches in their songs by listening to and interacting with adult males who are themselves skilled singers. This conclusion, in turn, led us to ask whether experience with males and their songs is necessary for chickadees to develop accurate relative-pitch perception. Isolated males sing only aberrant notes, but, we wondered, is this simply an impairment in relative-pitch production, or do the birds lack accurate relative-pitch perception?
To address this question, one of us (Weisman) and Milan Njevovan, now at the University of Alberta, housed both field-reared and isolation-reared chickadees individually in sound-resistant chambers and taught them to fly to a feeder to obtain food. Once a chickadee was standing on a perch directly opposite the feeder, we played a sequence of two pure-tone notes from a speaker beside the feeder. In this discrimination task, flying to the feeder after some two-note sequences was rewarded with food, whereas flying to the feeder after other two-note sequences was not rewarded.
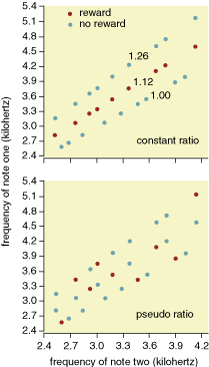
The logic of the experiment was that chickadees should be able to discriminate rewarded note sequences faster when they can categorize the rewarded note sequences by a common frequency ratio than when they need to memorize a random collection of rewarded note sequences one by one. Not surprisingly, when we played field-reared birds the note sequences with randomly rewarded frequency ratios, they needed twice as many sessions to acquire the discrimination as field-reared birds that were in the constant-ratio group. Most important, isolation-reared birds needed four times as many sessions as field-reared birds to learn the constant-ratio discrimination. It appears that field-reared male chickadees can perceive the constant ratio in the rewarded note sequences, whereas isolation-reared males are impaired in relative-pitch perception and instead need to memorize the rewarded note sequences one by one. Extrapolating from our results to the perception of fee-bee songs, we can conclude, first, that free-living male chickadees perceive but seem to ignore the constant internote ratio in rivals' songs during territorial interactions and, second, that males reared in isolation have difficulty in perceiving, as well as in producing, the internote relative pitch change in fee-bee songs.
We wondered about the possible evolutionary benefits of male chickadees' ability to shift pitch and female chickadees' knack for identifying pitch changes. In what follows, it may seem that we have granted chickadees remarkable foresight and cognitive powers to choose ways of behaving that will maximize their reproductive success. In fact, we are only using a shorthand common among biologists to describe the way selection has favored behaviors that result in greater reproductive success.
Chickadees are only one of several songbirds that transpose their songs over a wide range of frequencies. For species with a single type of song, transposing a song to match a rival's pitch might be an aggressive act, similar to singing the same song type as a rival in a species with multiple song types. Over most of North America, male black-capped chickadees sing a single song type, the fee-bee song. One of us (Ratcliffe) in collaboration with Dan Mennill, now at the University of Windsor, found that in duels over territory, male chickadees indeed use pitch matching to escalate conflicts with their rivals. Maintaining fixed frequency ratios in song might be one way chickadees provide consistent cues to their identity across song transpositions. But given that males do not seem to discriminate particularly among songs with varying internote pitch intervals, we concluded that the adaptive benefits of pitch matching must lie elsewhere.
We turned to the dawn chorus, that critical time of day when, among many songbirds, males advertise for mates, and females, even those with partners, prospect for mating opportunities. (Mating outside the pair bond is called extra-pair copulation.) Male chickadees regularly shift their songs during dawn singing, when overt territorial conflicts are rare. This suggests the motivation for frequent pitch transpositions during dawn singing might be communication with females.
This led Ratcliffe and former graduate students Peter Christie and Dan Mennill to study singing during the dawn chorus in more detail. We suspected that males might be shifting pitch to convey information about their quality to females, information females might value. Female chickadees prefer high-ranking males as social mates, with rank referring to dominance position in the previous winter's flocks. If females are relegated to a lower ranking social mate, they choose high-ranking males as partners in extra-pair copulations. Such matings account for the paternity of young in as many as a third of their clutches, as revealed by genotype analysis of the mother, the young and the social mate.
Male song can divulge the developmental conditions, genetic quality and current vigor of the singer and is known to affect females' choices in social mates and copulation partners. We measured the social rank of chickadees during aggressive encounters at winter feeders where high-quality males prevail, then compared these dominance rankings with several fee-bee song features recorded during dawn singing. The most important finding was that across multiple pitch transpositions, high-ranking males maintain the internote frequency ratio with much lower variability than do lower-ranking males. In musical terms, when higher ranked males transpose their songs upward in pitch during dawn singing, they hold the interval—that is, keep the internote ratio constant—whereas lower ranking males sing sharp—that is, they reduce the frequency ratio.
Previously, we found that female chickadees appreciate the quality of male songs in laboratory tests, but do female chickadees really listen to song duels in the wild? Ratcliffe together with Peter Boag, also of Queen's University, and Dan Mennill conducted a striking test that showed females do indeed eavesdrop on their mates' encounters with rivals. In 6-minute playback tests, we played recordings of high-ranking males having simulated encounters with an intruder. The mock-intruder either sang submissively, avoiding matching the resident's pitch, or aggressively, pitch matching the resident. The results showed that high-ranking males that lost in the simulated encounter with an aggressive intruder also lost paternity. Extra-pair paternity in their mates' clutches rose from 10 percent to 50 percent after submissive and aggressive playback respectively. Evidently, the female social partners of the recorded males engaged in more extra-pair copulations when they perceived a change for the worse in the quality of their social mates. These results suggest male chickadees have good reason to advertise their quality by holding constant the internote frequency ratio in their songs—female chickadees are listening carefully.
When we began studying songbirds almost twenty years ago, we chose black-capped chickadees because they were abundant and sang simple songs. Subsequently, our work has revealed that chickadees' seemingly simple songs present precise, discrete relative pitch cues. Males shift the starting pitch of their songs to match a rival's song as a threat and to display their vocal virtuosity to females during the dawn chorus. By preserving the relative-pitch relations between notes during these shifts, males provide reliable clues about their identity and quality.
Music has been part of the human experience since our inception as a species. Like chickadees, young humans learn about relative pitch from adult humans. Humans use precise, discrete pitch changes in music and transpose melodies while keeping the intervals between the notes constant. As among songbirds, humans tend to find musicians sexually attractive. Under the circumstances, it seems remarkably easy to relate to black-capped chickadees. It appears likely that at least several, if not most or all, oscine species can perceive relative pitch changes. As a resource, relative pitch may be ignored by some songbirds and used by others, but it is certainly exploited relentlessly by black-capped chickadees.
Click "American Scientist" to access home page
American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.