Exercise Controls Gene Expression
By Frank Booth
The activity level of skeletal muscle modulates a range of genes that produce dramatic molecular changes—and keep us healthy
The activity level of skeletal muscle modulates a range of genes that produce dramatic molecular changes—and keep us healthy

DOI: 10.1511/2005.51.28
Consider what likely would happen in the game show Jeopardy if the category what tissues do were to roll up on the game board. The $200 answer: "It moves the body." The contestant delivers the question—"What is Muscle?"—earns her money, and the game quickly moves on. This scenario typifies how muscle is generally viewed—as a simple part of the human anatomy that locomotes us.
Perhaps late-night television would allow a more sophisticated treatment of muscle's function. Suppose David Letterman were to ask people on the street "What is skeletal muscle?" Some of the answers might be: "What I see at the meat counter"; "what I feel the day after those rare occasions that I exercise"; and "what I saw on TV during the Olympics."

Jacques Langevin/Corbis Sygma
All these responses in fact vastly underplay the role of muscle, even the subcategory of skeletal muscle, in our bodies. In the following paragraphs, we hope to present a more sophisticated view of the function of skeletal muscle from a genetic and molecular standpoint, and in particular to emphasize the role of skeletal muscle in human health and well-being. Let's start by recognizing that skeletal muscle is the largest single tissue type in the human body: We have more than 640 muscles, accounting for between 30 and 40 percent of total body weight. Skeletal muscle also uses as much as 25 percent of the energy consumed by the body at rest.
Furthermore, although today we consider Olympic athletes to be extraordinarily fit, most of our ancestors from tens of thousands of years ago had bodies and muscles that were almost as fit. Many of the metabolic characteristics of modern humans evolved to support high levels of work efficiency and physical activity, a fact that undoubtedly contributes to the health problems facing sedentary 21st-century societies. Our underuse of skeletal muscle may play an underrecognized role in the rise of chronic diseases as a cause of modern mortality. Given these few simple facts, it is clear that the importance of skeletal muscle to humans extends well beyond athletics.
Skeletal muscle continuously adjusts its composition to meet the acute or chronic demands placed on it—a process called plasticity. A very visual example of plasticity is what happens to skeletal muscle when the daily demand for weight-bearing activity is lost during bed rest, limb immobilization or space flight. One of us (Booth) has shown that one-third of skeletal muscle in an immobilized limb can disappear within weeks. It is as if skeletal muscle recognizes that it is not needed and "remodels" itself into weak muscle. The converse is also true: If the work demand on muscle increases in a sedentary individual, even for relatively short periods of time per day (for example, beginning an aerobic-exercise program), the muscle adaptively remodels its protein composition to allow energy to be used more efficiently when it contracts.
For example, Booth and others working in John Holloszy's laboratory at Washington University in St. Louis, and elsewhere, found that mitochondrial content and capillary density in rats nearly doubled in skeletal muscle as a consequence of a four-month progressive running program—and that these changes in protein composition were sufficient to allow the otherwise sedentary animals to become "marathoners." Inside the cells, mitochondria serve as skeletal muscle's power plant, using oxygen to make adenosine triphosphate (ATP) from breakdown products of glucose and fatty acids. Capillaries supply the needed oxygen to the mitochondria. A doubling of mitochondria and capillaries doubles the capacity of the muscle to make ATP with oxygen (known as aerobic capacity), and greater aerobic capacity allows the muscle to work longer before reaching the point of fatigue. How does this happen?
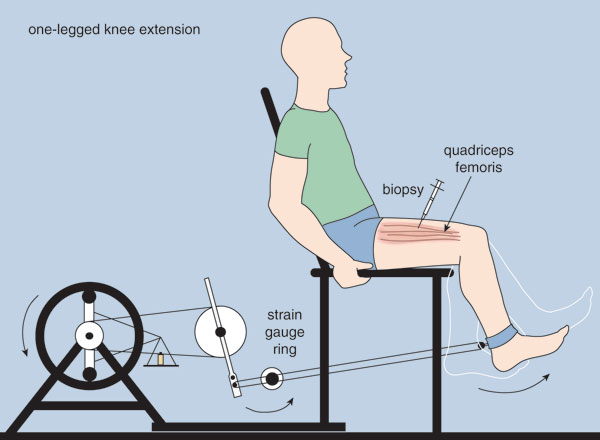
Illustration by Stephanie Freese, adapted from Andersen and Saltin 1985.
In order for skeletal muscle to exhibit plasticity, specific genes in the muscle sense the change in muscle usage and respond by altering the quantities of proteins that they produce. The mechanism by which genes in muscle undergo such profound responses during exercise training has recently been explored at the molecular level in humans. In collaboration with Bengt Saltin and Henriette Pilegaard at the Copenhagen Muscle Research Centre and George Ordway at the University of Texas Southwestern Medical Center at Dallas, one of us (Neufer) had subjects perform one-legged knee extension (kicking) exercise for 90 minutes per day for five consecutive days using a modified cycle ergometer (see Figure 2). In this study, the leg had to move a lever against a resistance that was about 70 percent of the maximal effort the subject could achieve for two minutes. On the fifth day of training, muscle biopsy samples were obtained from the untrained leg before exercise (control), and from the trained leg before and immediately after the 90-minute exercise-training session and at various times during the subsequent 24-hour recovery period. The samples were then processed in Neufer's lab at the John B. Pierce Laboratory at Yale University to determine (by measuring messenger RNA concentration or how quickly a specific gene was transcribed) whether exercise training activated specific target genes, and whether the activation took place within a specific time frame.
Barbara Aulicino
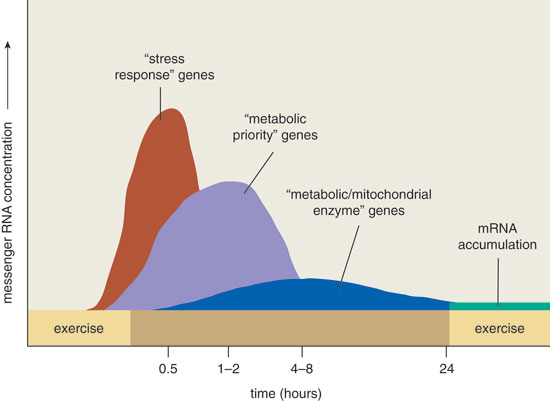
The results revealed that a large number of genes in the exercising skeletal muscle were activated after exercise (see Figure 3). These genes can be broadly divided into three categories based on the duration of their activity and, to a lesser extent, the change in concentration of the proteins they produce. "Stress-response" genes are activated quickly during the later phases of endurance exercise. Their protein output rises to quite high levels and returns to normal levels very quickly after exercise ceases. These acutely and transiently sensitive exercise-responsive genes appear to encode proteins that are part of a general response to stress in all cell types. Proteins that fall in this category include heat shock proteins and some transcription factors (immediate early genes).
The "metabolic priority" genes, the second category, make proteins that are required as a consequence of a particular metabolic stress, such as when blood glucose or muscle glycogen levels become low. A select number of these genes were induced to express themselves in the exercised leg at 15 or more times the rate of induction in the untrained leg. Interestingly, the increases peaked during the initial few hours of recovery from the exercise bout but returned to control levels within 24 hours.
The third category, "metabolic/mitochondrial enzyme" genes, encode proteins whose function is to convert food to energy. These genes are activated much more slowly, reaching only modest increases over baseline. Nonetheless, their products persist for some time in tissue.
Stephanie Freese
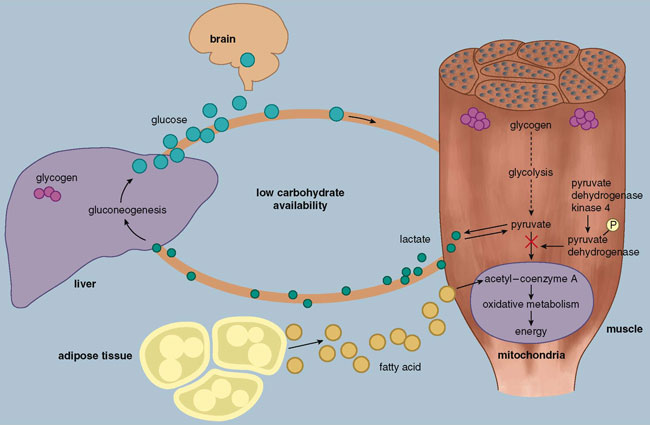
Let's look more closely at a few of the hundreds of genes modulated by muscle activity. The role of one of the "metabolic priority" genes might at first seem paradoxical. This gene codes for a protein known as pyruvate dehydrogenase kinase 4, or PDK4. When PDK4 appears in the cell, it inhibits the reaction that allows the breakdown products of glycolysis (glucose) to enter the mitochondria for further catalysis to ATP, thereby limiting the amount of energy gained from carbohydrates. Although shutting down a major source of energy for muscle during the later portions of prolonged aerobic exercise seems like a bad idea, consider that glucose is virtually the only fuel the brain can metabolize to make ATP (see Figure 4). When blood glucose availability becomes low (during prolonged exercise or fasting), the body cannot afford to allow skeletal muscle to continue to utilize glucose at the expense of the brain. Instead of entering the mitochondria, the breakdown products of glycolysis travel out of the muscle to the liver, where they are turned into new glucose molecules and released back into circulation to supply the brain. Thus the induction of PDK4 in skeletal muscle during the latter portion of a long exercise bout appears to serve as a natural defense mechanism reflecting priority given to the metabolic needs of the brain. Why then would PDK4 remain elevated or increase even further for several hours after exercise when the availability of glucose is no longer an issue? Although research is ongoing, here again the answer likely reflects a different priority after exercise: to ensure that glucose is being used to replenish glycogen stores (the storage form of glucose) in muscle and the liver as quickly as possible, preparing the body for the next challenge.
Neufer's findings also showed that the transcriptional activity of other genes, including the metabolic stress-related genes uncoupling protein-3 and heme oxygenase-1, increased by three- to sevenfold in response to exercise, peaking after one to two hours of recovery. That work, as well as several similar studies, further revealed that exercise transiently activates transcription of a large number of other mitochondrial genes but that the degree of activation is much smaller, increasing only two- or threefold (see Figure 3). The small transient rise was somewhat surprising at first, given that an increase in the expression of these genes is the response that ultimately leads to doubling of proteins in the mitochondria with a resultant improvement in metabolic function.
How could such small transient increases lead to the large increases in mitochondrial proteins associated with several weeks of endurance training? Again, although research is ongoing in this area, a key point is that mitochondrial enzymes typically have a slow turnover rate. For example, Booth showed that 50 percent of the mitochondrial protein cytochrome c in rat skeletal muscle is replaced every week. (Cytochrome c turnover contrasts sharply with that of the protein PDK4, which likely turns over within hours.) The relatively slow rate of replacement of old protein by new cytochrome c dictates that it takes one week for half of the final increase in mitochondrial protein levels to happen, a second week for half of the remaining 50 percent increase to a new, higher, steady state to occur, and so forth. It also means that even a slight increase in the concentration of cytochrome c will persist for several days to weeks because of the slow turnover of this protein. Thus the longevity of proteins that cause plasticity logically contrasts with the short lifetime of metabolic priority proteins such as PDK4, which are required only under certain specific conditions.
Collectively, exercise studies have led Neufer to hypothesize that the adaptations associated with several weeks of aerobic exercise training actually stem from the cumulative effects of the transient increases in gene expression that occur in response to each individual exercise bout. Only those proteins with a sufficiently long half-life will accumulate with successive bouts of exercise, independent of the level of activation of the corresponding gene. These findings explain why it takes weeks to months of aerobic training to improve the aerobic fitness of humans.
In contrast to the lengthy period needed to increase aerobic fitness, other metabolic changes take only a day or days to occur but then quickly disappear if daily physical activity does not continue. And although epidemiologists have long known that exercise is associated with lower blood sugar and insulin levels, recent investigations into the mechanism for this effect have produced remarkable results. Physical activity actually limits the rise in blood glucose after the next meal—and it does so after only a single bout of exercise!—by making insulin work better in moving glucose into muscle. In popular language, it is as though physical activity lowers the glycemic index of carbohydrates. Although research is ongoing in this area, it is thought that a single exercise session is able to accomplish this by affecting the property of one or more signaling proteins that are activated when insulin binds to the cell. These signaling proteins may then become more sensitive, leading to a more rapid movement of the GLUT4 glucose transporter protein to the plasma membrane (see Figure 6). In a resting muscle cell, GLUT4 resides in vesicles below the plasma membrane. In response to insulin or exercise, these GLUT4 vesicles fuse with the plasma membrane to transport glucose out of the blood and into the cell. In addition to exercise's stimulus to direct GLUT4 to the plasma membrane, Neufer has shown that repeated bouts of daily exercise activate the GLUT4 gene to make more GLUT4 mRNA and thus GLUT4 protein. Conversely, after a few days of physical inactivity in aerobically trained individuals, GLUT4 protein falls to levels found in the skeletal muscles of those patients with type-2 diabetes.
Body weight is controlled by the balance between the number of calories consumed and the number of calories expended. Caloric intake is determined solely by the amount and type of food ingested. Caloric expenditure is more complex as it is determined by four factors. The number of calories used each day is a function of the minimum energy required to keep cells, tissues and organs alive at rest (the basal metabolism, analogous to the idling of an automobile engine); the energy required for digesting food; the energy required to heat the body; and the energy required for movement or physical activity. The number of calories required for the first three is generally fixed from day to day in our modern world. Consequently, an individual's level of skeletal muscle usage has the largest influence of the four factors on day-to-day caloric balance and thus plays a key role in the cause of obesity.
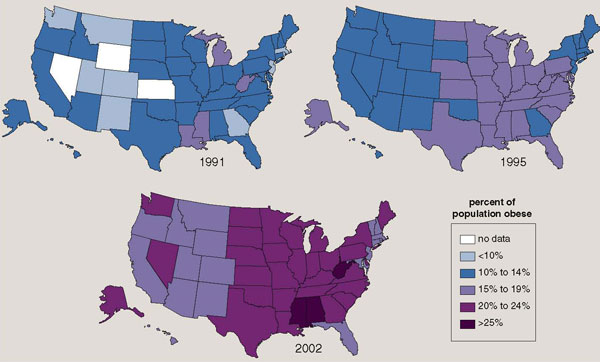
Illustration by Barbara Aulicino. Map data from the Centers for Disease Control and Prevention.
Does a reduction in physical activity lead to a gain in body fat? A simple example will demonstrate that the answer is "yes." A 70-kilogram man who reduced the distance walked by two football fields per day for a year would add a pound of fat if his caloric intake went unchanged. Subtract a kilometer (approximately the length of 11 football fields) from each day's walking for a year, and five additional pounds of fat would be gained. Obesity qualifies as an epidemic in the United States; currently more than 60 percent of adults are overweight and 30 percent are obese (see Figure 5). This is rapidly becoming a global problem; the World Health Organization estimates that 60 percent of people worldwide are overweight. Many health scientists view this disease as resulting primarily from overeating. They often overlook the fact that the average sedentary individual actually consumes fewer calories today than did his forebears a few generations ago.
Only a few centuries ago, human beings had to perform physical labor almost daily to survive. Acquiring food and shelter, defending against enemies and rearing offspring who would survive to reproductive age all required strenuous effort. Only in the past century did farms become sufficiently mechanized to eliminate much of the physical labor previously required by agriculture. Preparing materials for and constructing housing were done without power tools as recently as 100 years ago. Even transportation required more effort a century ago. Prior to the industrial and computer ages, the daily physical activity levels required just to get by were much higher than in today's mechanized, electronic society.
Although precise measurements of calorie usage during daily activity are not available from centuries ago, it has been estimated that the average hunter-gatherer expended about 1,000 kilocalories per day in physical activity while consuming about 3,000 kilocalories, a 3:1 ratio for what has been called subsistence efficiency. In contrast, the typical sedentary person in the 21st century consumes about 2,400 kilocalories per day but expends only about 300 kilocalories in physical activity, a subsistence efficiency ratio of 8:1. Thus, despite an overall reduction in caloric intake, the average person is burning a much smaller proportion of calories through physical activity on a daily basis and has a 100-kilocalorie-per-day excess. We suggest that a major cause of the obesity epidemic is the huge decline in physical activity brought on by the Industrial Revolution and the computer age. There is considerable empirical evidence that the fitness level of muscles affects the basal metabolic rate, but just how wasn't clear until recent investigations at the molecular level. Ron Evans of the Salk Institute engineered transgenic mice to overexpress the gene peroxisome proliferator-activated receptor delta (PPARδ). This caused an increase in skeletal muscle mitochondria, like the conversion of some muscle fibers to a slow-oxidative phenotype—the type developed by aerobic physical activity—which protected against the development of obesity, even without an increase in exercise. It turns out that the transgenic mice burned more calories when fed a high-fat diet than did mice without the extra protein from the PPARδ gene. Serge Luquet, then of Inserm U470 in France, found that exercise by mice increases the amount of PPARδ protein in skeletal muscle, producing a similar effect to that seen in Evans's transgenic mice. Fit mice and fit people use more energy than do sedentary individuals, even when they're not moving.
Tens of thousands of years ago, physical activity was not likely a luxury as it is now in developed countries. Those unable to move quickly and persistently were likely to perish before reaching reproductive age, preventing their genes from being perpetuated. Has human evolution actually selected for genes that promote physical activity? The answer to that provocative question may be "yes." Evidence suggests that the metabolic status of skeletal muscle may influence not only the ability but also the will to run distances.
When Maureen Charron at Albert Einstein College of Medicine and her collaborators genetically engineered mice to make an excess of the protein GLUT4 in their skeletal muscle, the mice voluntarily ran four times farther each day than did normal mice. These results can be interpreted to suggest that the brain senses the potential of skeletal muscle to remove glucose from the blood, the function of GLUT4.
A different set of experiments show that the higher the aerobic capacity of skeletal muscle, the longer the distance that rats can and do run. Thirty years ago, Robert Fitts in John Holloszy's lab reported that exercise training produced a direct correlation of 0.79 between the increased expression of the cytochrome c protein and the ability of rats to run long distances. Cytochrome c is located in the electron-transport chain of mitochondria, where it assists in converting energy from food to ATP. In this case, an increase in protein quantity allowed the interpretation that gene expression for cytochrome c was increased. Booth later verified that the cytochrome c gene could be responding to exercise when he observed, in addition to the protein turnover mentioned above, that cytochrome c messenger RNA increased in the skeletal muscle of rats who had aerobic training. These studies show that an increase in mitochondrial proteins such as cytochrome c is associated with longer bouts of aerobic activity, but a more causal relationship has recently been found.
Back at the Salk Institute, the same group of mice that had their skeletal muscle genetically engineered to overexpress PPARδ had an increase in mitochondrial protein volume in their skeletal muscle. When tested to see how far they could run on treadmills, the genetically engineered mice were able to run almost twice as far as their normal relatives. Thus, enhancing aerobic enzymes in skeletal muscle increases the ability to exercise. The health of the aerobic machinery in skeletal muscle appears to determine how much aerobic exercise may be performed.
As evidence mounts that selective pressure has favored genes promoting physical activity throughout most of our species' existence, it becomes clear that many metabolic characteristics in modern humans likely evolved tens of thousands of years ago. This poses a paradox.
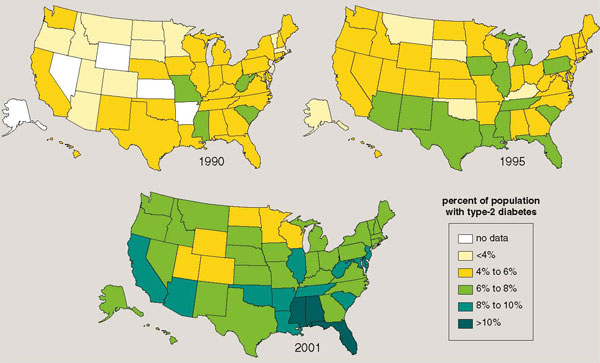
Illustration by Barbara Aulicino. Map data from the Centers for Disease Control and Prevention.
The same genes that were selected for survival when people had to forage for their food thousands of years ago may now shorten survival in the absence of sufficient physical activity. In the inactive individual, these genes make different quantities of proteins that interact with proteins from susceptibility genes for chronic diseases. Over time, these interactions can reach a threshold of biological significance that results in overt clinical disease. Atherosclerosis, certain cancers, obesity, type-2 diabetes, weakness in old age and many other chronic diseases are increased by sedentary lifestyles. Diagnosis of type-2 diabetes at the age of 40 years shortens the average lifespan of males and females by 12 and 14 years, respectively. Failure to use the genes for physical activity decreases survival. As an example, let's look at how physical inactivity contributes to the progression to type-2 diabetes.
The effect of physical activity and inactivity on insulin's action to remove blood glucose may have been selected as a protective process during evolution. The human body stores very little glucose in muscles and the liver (it becomes depleted within 24 hours of fasting). Therefore, it needs mechanisms to be stingy in its use of glucose when body stores are near empty—-such as during famine or after prolonged exercise. (Recall that PDK4 deprives muscles of glucose to protect the brain.) Consequently, it would have been advantageous to our ancestors to develop a system that allowed only physically active muscles to remove blood glucose. We posit that just such a mechanism is in evidence when only working muscle activates GLUT4 to transport blood glucose into the muscle. We speculate that skeletal muscle has been engineered through evolution to have physical activity and inactivity cause and prevent, respectively, glucose uptake into skeletal muscle. Although this was an exquisite system for survival 10,000 years ago, a continuously sedentary life-style today keeps blood glucose and insulin levels high, leading to type-2 diabetes. The best way to reverse the high levels of blood insulin and glucose in sedentary individuals is to have muscles contract during exercise.
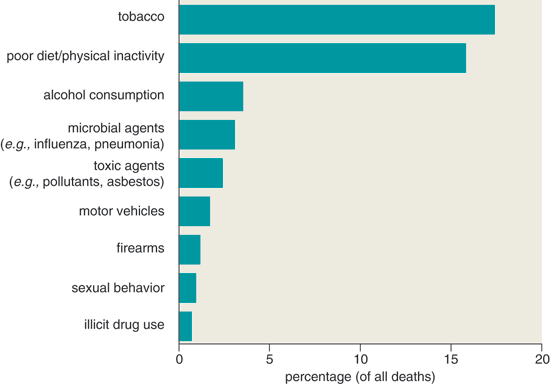
Barbara Aulicino
To emphasize just how profound this effect is and how recently it has become critical, consider that a century ago type-2 diabetes was never seen in humans younger than 40 years. Even two decades ago it was routinely called "adult-onset diabetes." Today, physicians are seeing "adult" diabetes in 10-year-olds. But it's not just diabetes that's cropping up in youth. Sixty percent of overweight teenagers already have at least one risk factor for coronary artery disease. Whatever merits youth sports may have for social development, young people (and old) need physical activity outside of sports just to maintain metabolic homeostasis to prevent many chronic diseases that will shorten their lives.
Evidence from epidemiological studies shows that a sedentary life-style increases the incidence of more than 20 chronic diseases. For example, among 70,000 female nurses aged 40 to 65 years in the Nurses' Health Study, when nurses who walked the equivalent of three or more hours per week at a brisk pace had 30 percent less coronary artery disease, ischemic stroke and type-2 diabetes compared to sedentary nurses. Some site-specific cancers are also more prevalent when moderately active females become inactive. Breast cancer rates rose 25 percent among the sedentary in one study, and each one-hour decrease of recreational physical activity per week during adolescence was associated with a 3 percent increased risk of breast cancer. In a Scandinavian study of twins, life-style caused 58 to 100 percent of sporadic cancers. Such research underlies the recent decision by the U.S. Centers for Disease Control and Prevention (CDC) to designate sedentary life-style as an actual cause of most chronic diseases (see Figure 7). Indeed, physical inactivity is now the third leading cause of death in the United States and contributes—to a great extent, we argue—to the second leading cause (obesity). Physical inactivity is the cause of least 1 in 8 deaths.
Lack of exercise is only one of many life-styles killing people in affluent nations. The major three current life-style factors (tobacco, poor nutrition and physical inactivity) account for 1 in 3 deaths in the U.S. However, other data place this percentage higher. Ninety-one percent of the cases of type-2 diabetes and 82 percent of the coronary artery disease cases in 84,000 female nurses in the Nurses' Health Study were attributed to unhealthful habits and so-called high-risk behaviors. (A high-risk life-style is defined in the study cited as a body mass index of greater than 25, a diet low in cereal fiber and polyunsaturated fat and high in trans-saturated fat and glycemic load, a sedentary life-style and current smoking.) Obese males and females live an average of 6 and 7 years less, respectively, than their trim counterparts.
It is also worth emphasizing that deadly life-styles that produce chronic diseases are a relatively recent development. Since 1900 there has been a dramatic shift from a preponderance of infectious disease to a preponderance of chronic disease affecting both morbidity and mortality. As a society, we have not adapted well to this change. We ignore not only the prevention but also the progression of chronic diseases. Even if we take action before the diseases prove fatal, full recovery is often impossible.
Is the increase in chronic diseases all of genetic origin? The answer is both "no" and "yes." According to the CDC, the answer is "no" if we are referring to disease-susceptibility genes; there's no evidence that we are becoming more genetically susceptible to heart disease, cancer or other chronic diseases. According to us, the answer is "yes" if we are referring to genes whose expression is altered by life-style. For example, the GLUT4 gene is in reality a life-style gene. Its expression is altered by the level of muscle contraction. The failure to be active enough to have GLUT4 in the plasma membrane of skeletal muscle predisposes one to be pre-diabetic.
Finally, having weak skeletal muscles has been shown to be a health threat in and of itself, at least in the elderly. The prevalence of clinically significant weak skeletal muscle is estimated to range from 8.8 percent in individuals less than 70 years old to 17.5 percent in those 80 and older, and recent research indicates that these people are at greater risk for mortality compared with age-matched individuals without weak skeletal muscles. Further, as the amount and the rate of loss in size of skeletal muscle increases, the risk of premature death also increases. John Morley of St. Louis University and others have written that decreased physical activity with aging appears to be the key factor involved in producing weak muscles in old age. A vicious cycle exists between physical inactivity and muscle strength. Weaker and smaller skeletal muscles with aging engender reduced physical activity, the diminished physical activity further decreases the size of skeletal muscle, and the downward spiral continues.
Work to understand how physical inactivity changes the mixture of proteins made as old skeletal muscle becomes physically inactive remains ongoing. Booth and Scott Pattison of the University of Missouri used small glass plates containing thousands of copies of genes to measure mRNA made by muscle. We found that more than 700 genes (out of about 24,000 measured) changed when comparing old to young skeletal muscles. Since many patients with, for example, broken hips successfully survive the surgical repair but later die with weak skeletal muscles, we wanted to identify the genes responsible for the inability of the weak skeletal muscles to get strong again in old humans. Old rats experience a similar loss in ability to enlarge skeletal muscle after limb immobilization, so we looked at differences using the animal model. A total of 354 genes differed in their expression in skeletal muscle between young and old rats with immobilized limbs. As we write, our current work focuses on determining which of these genes are the culprits that prevent recovery of skeletal-muscle strength after limb immobilization.
By now we hope you'll agree that skeletal muscle is a fascinating and largely underrated tissue. But a richer scientific understanding of its function may also require some changes in the linguistic world. The profound effects of underutilizing skeletal muscle suggest that the old adage "use it or lose it" turns out to be a gross understatement.
The authors gratefully acknowledge support from the National Institute of Arthritis and Musculoskeletal and Skin Diseases at the National Institutes of Health.
Click "American Scientist" to access home page
American Scientist Comments and Discussion
To discuss our articles or comment on them, please share them and tag American Scientist on social media platforms. Here are links to our profiles on Twitter, Facebook, and LinkedIn.
If we re-share your post, we will moderate comments/discussion following our comments policy.